BioAcyl Corp |
|
Newson, J., Stables, M., Karra, E., & others. (2014). Resolution of acute inflammation bridges the gap between innate and adaptive immunity. Blood, 124(11), 1748–1764. Added by: Dr. Enrique Feoli (12/02/2021, 23:25) Last edited by: Dr. Enrique Feoli (10/05/2023, 17:59) |
| Resource type: Journal Article DOI: 10.1182/blood-2014-03-562710 ID no. (ISBN etc.): 0006-4971 BibTeX citation key: Newson2014 View all bibliographic details |
Categories: BioAcyl Corp, BioAcyl Corp Subcategories: Inflammation Post resolution, Inflammation resolution Creators: Karra, Newson, others, Stables Collection: Blood |
Views: 3/284
|
| Abstract |
|
Acute inflammation is traditionally characterized by polymorphonuclear leukocytes (PMN) influx followed by phagocytosing macrophage (Mφs) that clear injurious stimuli leading to resolution and tissue homeostasis. However, using the peritoneal cavity, we found that although innate immune-mediated responses to low-dose zymosan or bacteria resolve within days, these stimuli, but not hyperinflammatory stimuli, trigger a previously overlooked second wave of leukocyte influx into tissues that persists for weeks. These cells comprise distinct populations of tissue-resident Mφs (resMφs), Ly6chi monocyte-derived Mφs (moMφs), monocyte-derived dendritic cells (moDCs), and myeloid-derived suppressor cells (MDSCs). Postresolution mononuclear phagocytes were observed alongside lymph node expansion and increased numbers of blood and peritoneal memory T and B lymphocytes. The resMφs and moMφs triggered FoxP3 expression within CD4 cells, whereas moDCs drive T-cell proliferation. The resMφs preferentially clear apoptotic PMNs and migrate to lymph nodes to bring about their contraction in an inducible nitric oxide synthase-dependent manner. Finally, moMφs remain in tissues for months postresolution, alongside altered numbers of T cells collectively dictating the magnitude of subsequent acute inflammatory reactions. These data challenge the prevailing idea that resolution leads back to homeostasis and asserts that resolution acts as a bridge between innate and adaptive immunity, as well as tissue reprogramming.
|
| Notes |
DiscussionWe show that there is a previously overlooked third phase of leukocyte influx into tissues after onset and resolution of acute inflammation. These cells comprise Ly6chi-derived moMφs, moDCs, and MDSCs. In addition, tissue-resident (prenatal-derived17-19 ) resMφs that disappear during the early phase of the inflammatory response,20 reappear postresolution. These diverse populations of mononuclear phagocytes were observed alongside lymph node expansion and increased numbers of peripheral blood and peritoneal memory and regulatory lymphocytes. Based on our data and supported by others,21-24 we conclude that in response to resolving, but not hyperinflammation, DCs residing in naïve tissues take up antigens and migrate to local lymph nodes (including peritoneal milky spots25 ) to initiate lymphocyte activation. The latter is amplified by postresolution CCR2-expressing monocytes; whether these effector monocytes26 exert their effects after migrating into the cavity and/or directly from the blood21 remains to be clarified. Concomitantly, as inflammation resolves resMφs and moMφs trigger FoxP3 expression within CD4 T cells while moDCs trigger their proliferation. The resMφs repopulate the cavity with a proportion migrating to the draining lymph nodes to bring about lymph node contraction in an iNOS-dependent manner from day 9 to day 13 onward. Finally, populations of moMφs remain in tissues months after the inflammation has resolved, dictating the magnitude of subsequent acute innate inflammatory stimulation (see hypothesis shown in supplemental Figure 7). The importance of IL-10 and TGF-β in zymosan-generated Mφs, in terms of counterbalancing adaptive immune responses, was reported by others.27 The resMφs expressing TGF-β and IL-10 as a consequence of phagocytosing apoptotic PMNs28 migrate to the lymph node and spleen expressing immune-suppressive iNOS. These cells are likely to remain in lymphoid organs with a role in terminating adaptive immune responses and long–term, tempering of the severity of future antigen-specific immunity. A role for iNOS in this setting was illustrated in the context of “adjuvant immunogenicity,” first described more than 40 years ago, which was found to be dependent on the presence of Mycobacterium tuberculosis within the adjuvant.29 Exposure to complete Freund’s adjuvant can impair the subsequent expression of autoimmune disease in rodents. This immunoprotective effect was demonstrated in multiple autoimmune disease models, both spontaneous and induced and in multiple species, including rats,30-33 mice,34 and guinea pigs.29,35 In each case, preimmunization with Freund’s complete adjuvant alone up to a month before the disease induction by immunization resulted in decreased incidence and severity of disease29-31,35,36 in an iNOS-dependent manner.37 Similarly, Mφ-mediated immunosuppression has been reported after bacteria, fungi, and parasite38 infection. For instance, mice immunized with attenuated Salmonella typhimurium (SL3235), although protected against virulent challenge, are unable to mount in vivo and in vitro antibody responses to non-Salmonella antigens, such as tetanus toxoid and sheep red blood cells, and exhibit profoundly suppressed responses to B- and T-cell mitogens. It transpires that suppression of antibody responses is mediated by iNOS within Mφs.39-42 Collectively, we argue that, in addition to bridging the gap between innate and adaptive immunity, resolution may also establish a phase of immunologic tolerance. It is unclear why a PMN-driven, acute onset phase of inflammation should be accompanied, paradoxically, by a prolonged phase of immune suppression. One possibility is to suppress the development of maladaptive immune response leading to autoimmunity. In contrast, injecting high-dose zymosan (10 mg) resulted in a substantial inflammatory cell infiltrate comprising classically activated (M1)-like Mφs secreting high levels of TNF-α, IFN-γ, and IL-6 (>2000 pg/mL). Compared with resolving inflammation, such an inflammatory insult triggered substantially reduced numbers of Tregs and effector/memory lymphocytes. Proinflammatory cytokines, such as interferon (IFN) and TNF-α, interfere with antigen-specific T-cell responses43-45 and clearance of viral and mycobacterial infections in mice.46,47 Furthermore, inhibition of these cytokines enhances pathogen clearance and resolution of disease, an effect that is dependent on the presence of T cells. This suggests that excessive inflammation inhibits antigen-specific T-cell function and therefore immunity to pathogens. Although the M1/M2 classification of macrophages was largely borne out of isolated monocytes incubated with defined growth factors in vitro, the phenotype of Mφ populations is likely to be more complex and overlapping, contingent on tissue, phase of inflammation, and the nature of the inciting inflammatory stimuli. We identified at least 4 distinct populations of monocyte/Mφs, each with a role in the resolution and in the development and control of adaptive immunity. In terms of individual cell phenotypes, we found that resMφs expressed TGF-β1 and IL-10, presumably as a consequence of phagocytising apoptotic PMNs; these cells also expressed ALOX-15 and TIMD4 to facilitate the recognition and uptake of apoptotic cells.12-14 In contrast, moMφs expressed IL12p35, whereas moDCs were enriched for IL1β, CCR7, CCR2, and CCL7. Taking this further, populations of moMφs that persisted in the cavity for months postinflammation were phenotypically different from resMφs. These data underline the fact that despite experiencing the same inflammatory cues, different monocyte/macrophage populations possess diverse phenotypes that are neither M1 nor M2, but are commensurate with the phase of inflammation. Indeed, this probably extends to macrophage populations occupying different tissue niches under physiological48,49 and disease conditions. As reported by others using thioglycollate-induced peritonitis,17 we also found a population of moMφs that persisted in the peritoneum for at least 2 months postresolution. The phenotype of these cells was different to that of resMφs at this time; indeed the phenotype of day 60 moMφs was also different than the phenotype early in the inflammatory response (at 72 hours). Moreover, the phenotype of resMφs 60 days post-zymosan was different to the inflammatory characteristics in the naïve cavity. This emphasizes functional plasticity in macrophage phenotype congruent with the environment. Specifically, despite coexisting in the same inflammatory milieu, macrophages of different origins acquire distinct phenotypes that change throughout inflammation. This emphasizes that far from revering back to the state, the tissue experience before resolution, postresolution tissues experience a state of “adaptive homeostasis,” which dictates the magnitude of subsequent inflammatory stimuli. This is an area that requires further exploration in the future. In summary, we show that resolution is not the end of the immune response to infection/injury but that it acts as a bridge between innate and adaptive immunity, thereby adding a third phase to acute inflammation after acute and resolution, namely postresolution. Disruption of proresolution pathways by hyperinflammatory stimuli, for instance, impairs the development of specific immunity. These data redefine resolution as the creation of a tissue microenvironment that facilitates interaction between the innate and adaptive arms of the immune system and that postresolution tissue that acquires a state of “adapted homeostasis.” |
| Quotes |
|
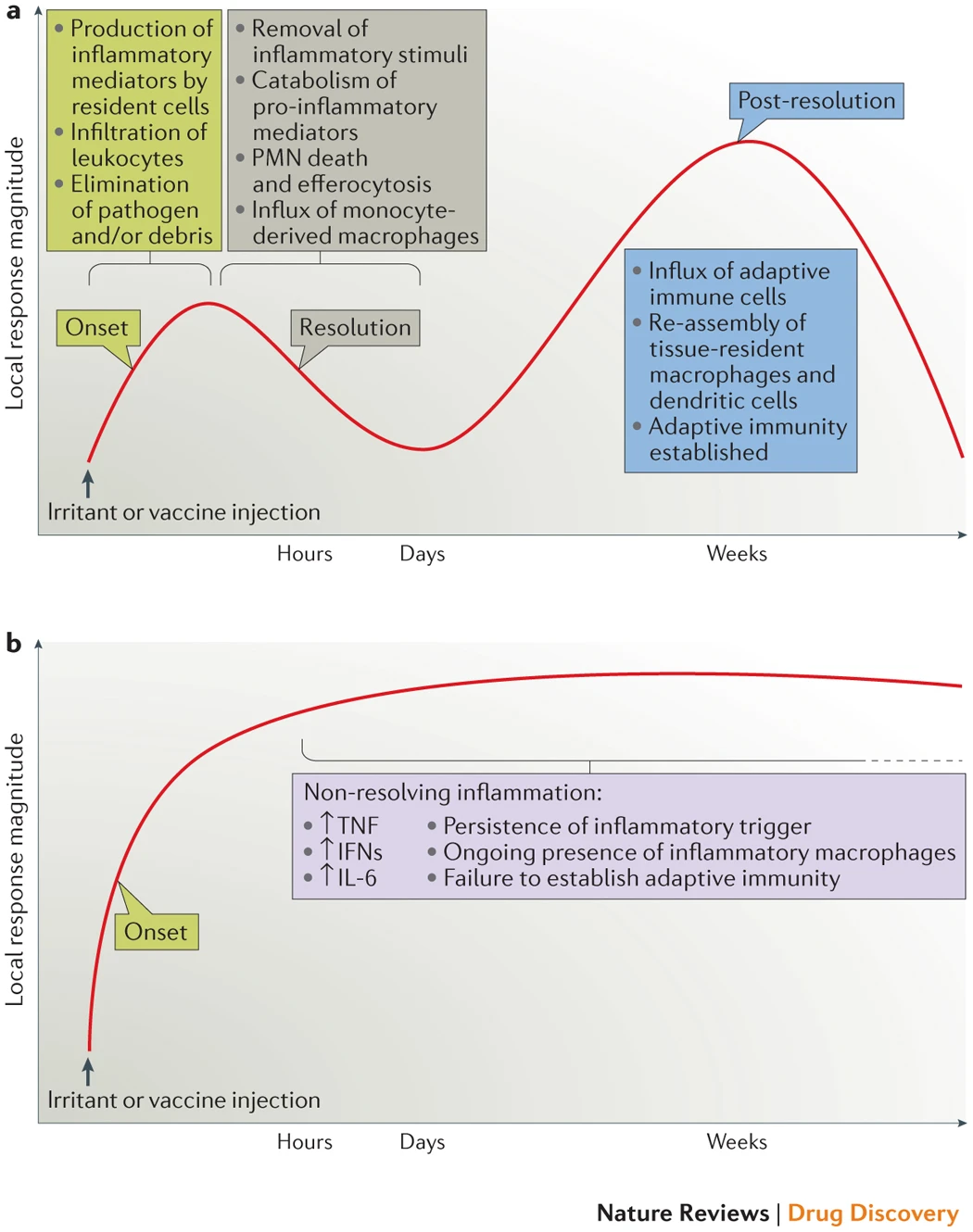
We have recently shown4 that resolution is not the end of the immune response to infection or injury, but rather that it acts as a bridge between innate and adaptive immunity, thereby adding a third phase after acute inflammation and resolution — namely, post-resolution (part a). Moreover, the idea that acute inflammation drives adaptive immunity also leads to an alternative explanation of the pathogenesis of some diseases that are driven by chronic inflammation (part b). Specifically, diseases driven by 'inflammation gone wrong' may arise from incomplete resolution of the initial acute response that, in turn, does not fully engage an appropriate adaptive immune response that would otherwise lead to full resolution. Thus, although many early-phase T helper 1-type cytokines have evolved to drive inflammation, their persistence may paradoxically derail resolution, resulting in impaired antigen clearance and the development of maladaptive immunity. IFN, interferon; IL-6, interleukin-6; PMN, polymorphonuclear cell; TNF, tumour necrosis factor.
Added by: Dr. Enrique Feoli
(2021-02-13 00:38:05)
|